
根據血流形態和位置的不同,流體剪切應力有不同的表現形式。例如,當血液在血管的直線部分流動時,會產生層流剪切應力(Lss),但當血液流過血管分支、分叉或彎曲時,會產生振蕩剪切應力或湍流。大量研究證實,動脈粥樣硬化(AS)在直血管中的發生率較低,而血管分叉處和彎曲處的動脈粥樣硬化發生率較高。因此,Lss 具有重要的抗 AS 作用。
CX3CL1 是由 373 個氨基酸組成的大分子蛋白,是目前已知的 CX3C 是趨化因子家族中的成員。研究報道,低剪切應力通過激活 CX3CL1 誘導單核細胞與 TNF-α(也稱為 TNF)激活的人臍靜脈內皮細胞(HUVECs)的粘附,而生理剪切應力消除了這種內皮炎癥。通過分析高通量測序數據,發現 CX3CL1 在 Lss 處理的內皮細胞中表達水平較低。因此,進一步的研究需要揭示 Lss 通過降低 CX3CL1 表達來抑制 AS 進展。
MicroRNAs(miRNAs)是一種長度約為 22 個核苷酸的非編碼單鏈 RNA,參與轉錄后基因表達的調控。研究提出,Lss 通過調節 miRNAs 來維持內皮細胞功能的穩定,從而發揮抗 AS 作用。有鑒于此,需要探討 Lss 敏感的 miR-29b-3p 對內皮細胞功能和 AS 形成的影響。
吉林大學人獸共患病研究教育部重點實驗室、新疆醫科大學基礎醫學院免疫學系、吉林大學第一醫院心血管疾病診治中心的聯合團隊的一項研究結合生物信息學、分子生物學和其他研究方法檢測了 Lss 敏感的 miR-29b-3p/CX3CL1 軸,在體外和體內探討了該軸對內皮細胞炎癥和 AS 發生發展的影響,從而提高了對 AS 調控的認識,為 AS 的防治提供了新的思路和見解。

為了探索 Lss 對血管內皮細胞功能的影響,實驗確定了在 Lss(12 dyn/cm2)處理的人主動脈內皮細胞(HAECs)和 HUVECs 中下調的 25 個 mRNAs 作為關鍵 mRNAs。
有研究報道,CXCR4、APLN、CX3CL1、CSF1 和 HES1 參與 AS 的發生和發展。因此,使用定量實時 PCR(qRT-PCR)來測量靜態和 Lss 處理的 HAECs 中 CXCR4、APLN、CX3CL1、CSF1 和 HES1 的 mRNA 表達。鑒于CX3CL1 mRNA 表達降低尤為顯著,于是將 CX3CL1 確定為后續研究的靶基因。與 HUVECs-static 組相比, HUVECs-Lss 組 CX3CL1 的 mRNA 表達也降低,與實驗中生物合成分析結果一致。
這表明,Lss 可以降低內皮細胞中 CX3CL1 mRNA 和蛋白表達,所以 CX3CL1 是 Lss 敏感基因。
研究報道,內皮 CX3CL1 誘導單核細胞粘附,從而破壞血管內皮功能。qRT-PCR 和蛋白質印跡顯示,與陰性對照組相比,TNF-α 誘導 HAECs 中 CX3CL1、血管細胞粘附分子-1(VCAM-1)和細胞間粘附分子-1(ICAM-1)的 mRNA 和蛋白表達增加。然而,在轉染 sh-CX3CL1-1 后,CX3CL1、VCAM-1 和 ICAM-1 的表達受到抑制(圖1 B-D)。此外,降低的 CX3CL1 表達抑制了 TNF-α 刺激的單核細胞粘附的增強(圖1 E、F)。
實驗進一步探討了 Lss 是否通過調節 CX3CL1 表達影響內皮細胞功能。HAECs 轉染 CX3CL1 過表達質粒(pri-CX3CL1)后,CX3CL1 的 mRNA 表達明顯上調,說明 CX3CL1 過表達細胞模型成功建立(圖1 G )。
如圖1 H-J 所示,Lss 顯著抑制 TNF-α 刺激的 CX3CL1、VCAM-1 和 ICAM-1 mRNA 和蛋白表達的增加,上調 CX3CL1 的表達逆轉了 Lss 的抑制作用。此外,對單核細胞粘附的分析表明,CX3CL1 的過表達部分逆轉了 Lss 對 TNF-α 刺激的單核細胞粘附增強的抑制作用(圖1 K、L)。
這表明,Lss 通過抑制 CX3CL1 表達損害單核細胞對活化 HAECs 的粘附。
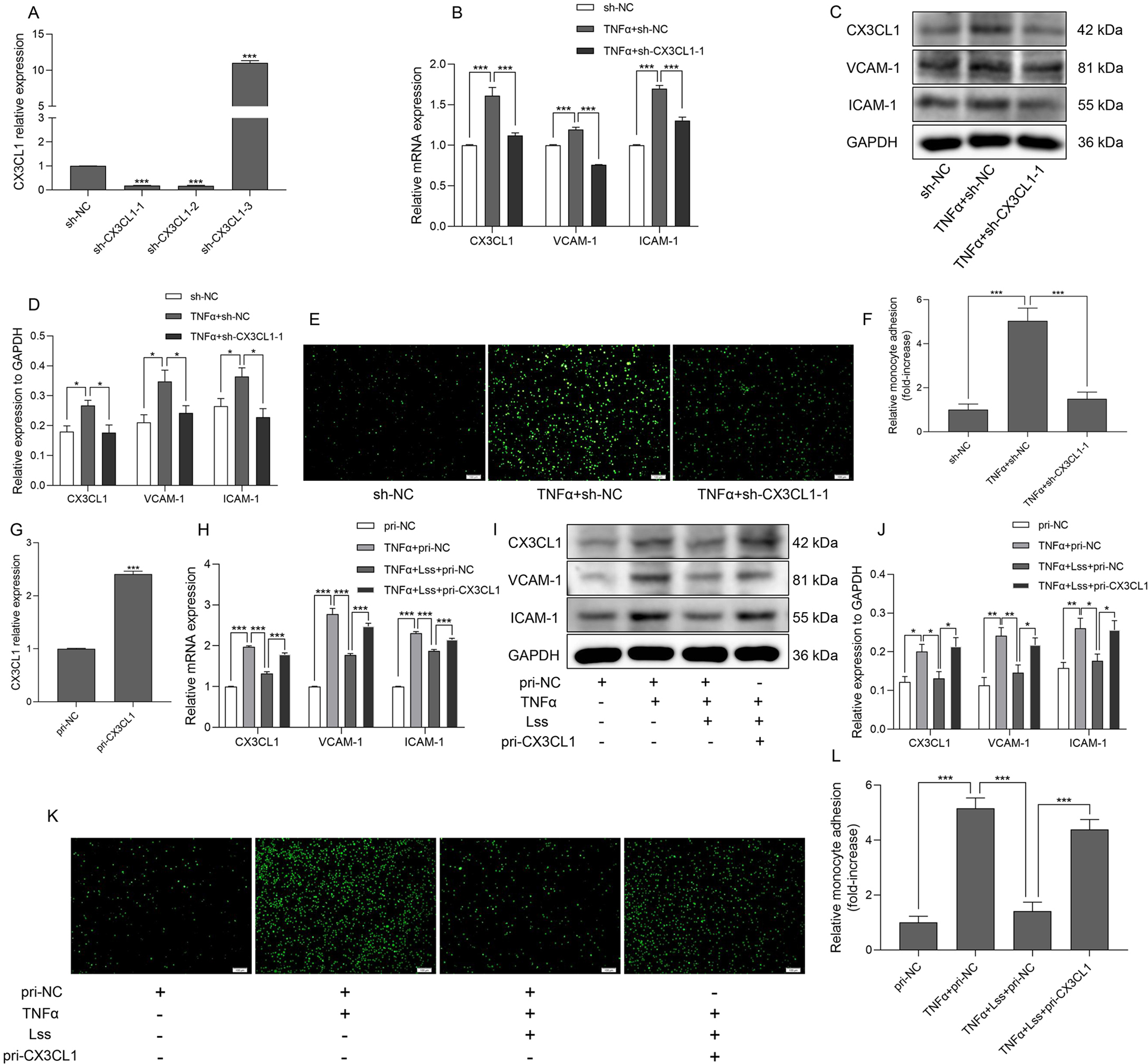
圖1 Lss 通過抑制 CX3CL1 的表達,損害單核細胞對活化 HAECs 的粘附。
最近的研究表明,受流體剪切應力調控的 miRNAs 是治療心血管疾病的潛在靶點。因此,為了分析 Lss 依賴性保護內皮細胞功能的分子機制,研究檢測了響應 Lss 調節 CX3CL1 表達的上游 miRNAs。
qRT-PCR 顯示,與 HAECs-static 組相比,Lss 尤為顯著上調了 miR-29b-3p 的表達。進一步驗證表明,過表達 miR-29b-3p 抑制 CX3CL1 mRNA 和蛋白表達,而敲低 miR-29b-3p 則促進 CX3CL1 mRNA 和蛋白表達。
這表明,CX3CL1 是 miR-29b-3p 的直接靶點。
隨后,研究探討了 miR-29b-3p 是否在 TNF-α 激活的 HAECs 中調節 CX3CL1 的表達。qRT-PCR 和蛋白質印跡顯示,miR-29b-3p 過表達顯著抑制 TNF-α 誘導的 CX3CL1 mRNA 和蛋白質表達升高(圖2 B-D)。此外,miR-29b-3p 過表達還顯著抑制 TNF-α 誘導的 VCAM-1 和 ICAM-1 mRNA 和蛋白質表達的增加,以及單核細胞粘附的增強(圖2 E-I)。
為了進一步驗證 miR-29b-3p 是否通過調節 CX3CL1 表達抑制單核細胞粘附到活化的 HAECs,將 pri-miR-29b-3p 和 pri-CX3CL1 共轉染到 TNF-α 激活的 HAECs 中。如圖2 J-L,在 TNF-α 激活的 HAECs 中,CX3CL1 的過表達消除了 miR-29b-3p 對 CX3CL1、VCAM-1 和 ICAM-1 mRNA 和蛋白質表達的抑制作用。此外,miR-29b-3p 過表達顯著抑制單核細胞與 TNF-α 激活的 HAECs 的粘附,而 CX3CL1 過表達逆轉了這種抑制作用(圖2 M、N)。
鑒于 miR-29b-3p 是 Lss 敏感的 miRNA,還探討了在加載 Lss 的條件下,miR-29b-3p 敲低對內皮細胞粘附的影響。如圖2 O-Q,在加載 Lss 后,TNF-α 上調的 CX3CL1、VCAM-1 和 ICAM-1 的 mRNA 和蛋白質表達受到抑制,而 miR-29b-3p 的敲低則逆轉了這種抑制作用。同樣,miR-29b-3p 敲低也逆轉了 Lss 對單核細胞粘附到 TNF-α 激活的 HAECs 的抑制作用(圖2 R、S)。
這表明,Lss 通過調控 miR-29b-3p/CX3CL1 軸損害單核細胞對活化的 HAECs 的粘附。

圖2 Lss 通過調控 miR-29b-3p/CX3CL1 軸損害單核細胞與活化的 HAECs 的粘附。在存在或不存在 TNF-α(10 ng/ml)的情況下,用 pri-miR-29b-3p 或陰性對照轉染 HAECs。
有研究提出,一些內皮 miRNAs 通過調節 NF-κB 信號通路來控制細胞粘附分子的轉錄,從而影響單核細胞與內皮細胞的粘附。為了進一步闡明 Lss 敏感 miR-29b-3p/CX3CL1 軸介導的內皮細胞粘附調節的分子機制,分析了 Lss、miR-29b-3p 和 CX3CL1 對 TNF-α 激活的 HAECs 中 NF-κB 信號通路的影響。
如圖3 A、B 所示,加載 Lss 后,TNF-α 誘導的 VCAM-1 和 ICAM-1 的上調受到抑制。此外,Lss 顯著抑制 TNF-α 誘導的 p65 和 IκBα(也稱為 RELA 和 NFKBIA)磷酸化(圖3 A、B),表明 Lss 可能通過抑制 NF-κB 信號通路的激活來調節內皮細胞炎癥。
p65 的核轉位是激活 NF-κB 信號的關鍵。如圖3 C-E 所示,加載 Lss 后,p65 在 TNF-α 激活的 HAECs 細胞核中的積累顯著減少。此外,免疫熒光分析表明,Lss 顯著降低了 TNF-α 誘導的 p65 核轉位(圖3 F)。值得注意的是,miR-29b-3p 敲低逆轉了 Lss 對 TNF-α 激活的 HAECs 中 NF-κB 信號通路的抑制作用。
實驗還探討了 Lss 敏感的 miR-29b-3p/CX3CL1 軸是否可以調節 TNF-α 激活的 HAECs 中的 NF-κB 信號傳導。如圖3 G、H 所示,miR-29b-3p 的過表達顯著抑制了 TNF-α 刺激的 VCAM-1 和 ICAM-1 的上調,以及 p65 和 IκBα 的磷酸化。在 CX3CL1 敲低的 TNF-α 激活的 HAECs 中獲得了相同的結果。miR-29b-3p 的過表達或 CX3CL1 的敲低減少了 p65 在 TNF-α 激活的 HAECs 的細胞核中的積累(圖3 I-K)。此外,免疫熒光分析還顯示,過表達 miR-29b-3p 或敲低 CX3CL1 顯著降低了 TNF-α 誘導的 p65 核轉位(圖3 L)。
這表明,Lss 敏感的 miR-29b-3p/CX3CL1 軸通過阻斷 NF-κB 信號通路減輕內皮細胞炎癥反應。

圖3 Lss 敏感的 miR-29b-3p/CX3CL1 軸通過阻斷 HAECs 中的 NF-κB 信號通路來減輕炎癥反應。
為了在體內將 Lss 與 miR-29b-3p 和 CX3CL1 聯系起來,實驗研究了暴露于湍流(Oss)的ApoE-/- 小鼠主動脈弓內膜和暴露于 Lss 的胸降主動脈內膜中 miR-29b-3p 和 CX3CL1 的表達水平。結果表明,與主動脈弓相比,暴露于 Lss 的胸主動脈內膜中 miR-29b-3p 的表達上調,而 CX3CL1 的表達下調。此外,體外研究發現,經振蕩剪切應力(0.5±4 dyn/cm2)處理的 HAECs 中 CX3CL1 的表達上調,而 miR-29b-3p 的表達下調。
為了確認 miR-29b-3p 是否在體內調節 AS 進展,給 6 周大的雄性ApoE-/- 小鼠喂食正常飲食(ND)或高脂飲食(HFD )4 個月。隨后檢測 ApoE-/- 小鼠血清中甘油三酯(TG)、總膽固醇(TC)、低密度脂蛋白膽固醇(LDL-C)、高密度脂蛋白膽固醇(HDL-C)的濃度。結果表明,與喂食ND的 ApoE-/- 小鼠相比,喂食 HFD 的 ApoE-/- 小鼠血清中 HDL-C 濃度顯著降低,而 TG、TC 和 LDL-C 濃度顯著升高。然而,當 miR-29b-3p 過表達時,HDL-C、TG、TC 和 LDL-C 的濃度恢復(圖4 C-F)。如圖4 G,與喂食 ND 的 ApoE-/- 小鼠相比,喂食 HFD 的 ApoE-/- 小鼠的主動脈內膜表面不光滑。伴隨著內膜增厚和較大的斑塊形成,表明 AS 小鼠模型建立成功。然而,當注射 agomir-miRNA-29b-3p 時,喂食 HFD 的 ApoE-/- 小鼠的主動脈內膜斑塊面積顯著減少(圖4 G )。
這表明,miR-29b-3p 在體內改善 AS。
實驗還探討了 miR-29b-3p 是否通過控制 CX3CL1 的表達來調節 AS 小鼠的內膜炎癥,從而影響 AS 的發生和發展。蛋白質印跡顯示,miR-29b-3p 的過表達逆轉了喂食 HFD 的ApoE-/- 小鼠主動脈內膜中 CX3CL1、VCAM-1 和 ICAM-1 蛋白表達的增加(圖4 J、K)。此外,免疫組化染色顯示,與喂食 ND 的ApoE-/- 小鼠相比,喂食 HFD 的 ApoE-/- 小鼠主動脈中 CX3CL1 和 VCAM-1 的表達顯著增加。然而,miR-29b-3p 的過表達下調了主動脈中 CX3CL1 和 VCAM-1 的表達(圖4 L)。
總之,miR-29b-3p 在體內減輕了內皮細胞炎癥,從而改善 AS。

圖4 MiR-29b-3p 在ApoE-/- 小鼠中減輕 HFD 誘導的內皮炎癥和 AS 。
總之,在喂食 HFD 的 ApoE-/- 小鼠中,Lss 敏感的 miR-29b-3p/CX3CL1 軸顯著抑制單核細胞與活化的 HAECs 的粘附并減輕局部炎癥和斑塊形成。因此,它是預防和治療 AS 的潛在干預靶點。
參考文獻:Pu L, Meng Q, Li S, Wang Y, Sun B, Liu B, Li F. Laminar shear stress alleviates monocyte adhesion and atherosclerosis development via miR-29b-3p/CX3CL1 axis regulation. J Cell Sci. 2022 Jul 15;135(14):jcs259696. doi: 10.1242/jcs.259696. Epub 2022 Jul 22. PMID: 35735031.
小編旨在分享、學習、交流生物科學等領域的研究進展。如有侵權或引文不當請聯系小編修正。
微信搜索公眾號“Naturethink”,學習更多細胞流體剪切應力技術及應用!
請輸入賬號
請輸入密碼
請輸驗證碼
以上信息由企業自行提供,信息內容的真實性、準確性和合法性由相關企業負責,化工儀器網對此不承擔任何保證責任。
溫馨提示:為規避購買風險,建議您在購買產品前務必確認供應商資質及產品質量。

 8
8
